

2020 Nov. 17 第9回“光”機到来!Qコロキウム
盛会のうちに幕を閉じました。ご参加ありがとうございました!
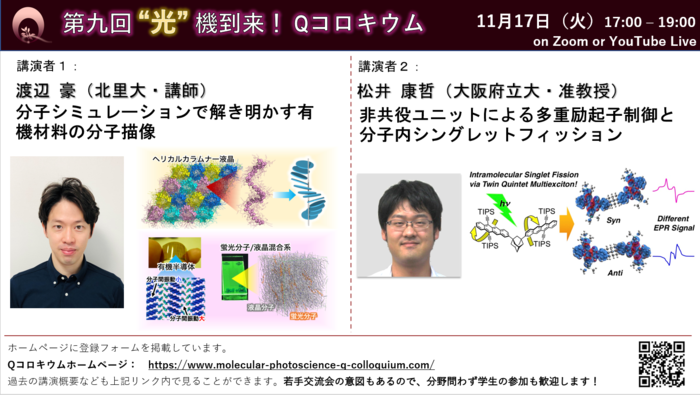
発表1:渡辺豪(北里大・講師)
分子シミュレーションで解き明かす有機材料の分子描像
有機材料は物理的なフレキシビリティーを失わずに特徴的な物性を発現するため、その応用の可能性は多岐に渡っており、様々な分野において機能性有機材料の開発が取り組まれている。しかしながら、その過程で必ずしも微視的観点からの構造や物性についての知見が得られているわけではない。我々は、分子レベルでの静的・動的構造を探る上で有用な分子動力学シミュレーションを用いて、様々な系においてミクロスコピックな描像の解明に取り組んでおり、本講演では最近得られた研究成果について紹介する。
Youtube チャットコメント一覧:
*チャットコメントに対する先生方からの返答はイタリックで追記しました。
質問① Kiyoshi Miyata
多分聞き逃したんですが、そもそもラジカルアニオンができるときの電子源はなんでしたでしょうか? 何にせよそこは律速になっていないということですよね。
質問② 山本 晃平
PITはある程度進行の進んだステージ4のがんなどに対しても有効なのでしょうか?
質問③ Kiyoshi Miyata
項間交差の量子収率そんなに低いんですね…代わりによく光ったりするんでしょうか?
質問④ Akio Yamauchi
軸配位の開裂は配位子2つのうちの片方しか起きていないようですが、一方の配位子に重原子のようなものを入れたりはできないのでしょうか?
質問⑤ Rie Wakabayashi
細胞膜上でスタッキングにより凝集が起こるとのことですが、抗体あたりにフタロシアニンは何分子くらい結合しているのでしょうか?
質問⑥ Kosuke Minamihata
細胞膜上のレセプターの数(結合した抗体の数)で毒性が変わるということですよね?抗体がアグリゲーションできるだけの十分な数が細胞膜上にないと減少が起きないと。
質問⑦ Akihisa Mino
誘導されるものがランダムな細胞膜損傷なのか、イオンチャンネルや他の受容体の活性化を介して浸透圧調整機構複合体や細胞骨格制御の上流にある受容体等が特異的に活性化されたりするのでしょうか。
普通の細胞にもがん細胞で発現される受容体を発現させると細胞死は誘導されるのでしょうか。
質問⑧ Youichi Tsuchiya
COTが三重項を吸った後はどうなるんでしょう?分解してラジカルが生成している可能性はありますでしょうか?
質問⑨ 笠 僚宏
PITは全身どこでも使えるのでしょうか。例えば脳腫瘍などに対しても有効なのでしょうか
質問⑩ Kiyoshi Miyata
項間交差の量子収率を測るのに蛍光相関分光を使われているのは何か理由があるのでしょうか? 過渡発光分光や過渡吸収分光で測れば時間スケールも含めて精密に評価できそうな気がします。
質問⑪ KEISUKE TANAKA
スタッキングは1抗体内で起こっている現象なのでしょうか?それとも凝集現象がドライビングフォースとなって膜内の複数の抗体をかき集めるような現象がおこっているのでしょうか?(レセプター自体が元から膜の一部へ集まっている?)
質問⑫ moriken55
もし細胞膜上で、すでに集まっているのでしたら、コンジュゲートは水中でも行くから会合しているのでしょうか。
質問⑬ moriken55
もし細胞膜上で、すでに集まっているのでしたら、コンジュゲートは水中でも会合しているのでしょうか。
コメント❶ moriken55
@南畑先生 膜タンパク質の膜上での動きは速いので、秒で毒性が出るのはありうると思います。
コメント❷ Kosuke Minamihata
森先生ありがとうございます。そうなのですね。
質問⑪への回答 Mikako Ogawa
KEISUKE TANAKA スタッキングは1抗体内で起こっている現象なのでしょうか?それとも凝集現象がドライビングフォースとなって膜内の複数の抗体をかき集めるような現象がおこっているのでしょうか?(レセプター自体が元から膜の一部へ集まっている?)
おそらく両方だと思います。近くにいる複数の抗体が集まることが効いていると思います。
発表2:松井康哲(大阪府立大・准教授)
非共役ユニットによる多重励起子制御と分子内シングレットフィッション
これまでに,優れた機能を有する数多くのπ共役分子が合成され,古くは染料や顔料,近年は有機エレクトロニクスなど,様々な分野で応用されてきた.一方で,π共役系の優れた機能をフルに引き出すには,非共役ユニットの効果的な利用により,共役系同士の相互作用を制御することも極めて重要である.本講演では,アダマンタンでTIPSテトラセンを連結した分子の”遅い分子内シングレットフィッション”を中心に,筆者の最近の研究を紹介する.
Youtube チャットコメント一覧:
質問① Tsuchiya Youichi
聞き逃したかもしれませんが、mgcTMPが細胞外膜ではなく内膜に局在する原理は何でしょうか?
質問② Rie Wakabayashi
基本的なところですが、一本鎖(Myr)では細胞膜親和性はなく、二本鎖になって初めて膜親和性が出るのは、リンカーの設計によるものでしょうか?
質問③ moriken55
ゴルジ体の外膜(?)にはアニオン性の脂質はあまりないのですか?膜を構成する脂質の組成を見る技術としても面白いのかもと思いました。
質問④ KEISUKE TANAKA
本筋から逸れるかもしれないのですが、1分子のmgcTMPからリンカー部を長く設定し複数に枝分かれさせて複数種のリガンドによる異種タンパク質の接近状態での局在化もしくは同一リガンド数を上げることでシグナル増幅などは可能なのでしょうか?内部反応経路に詳しくないため単純な話ではないかもしれませんが、興味本位で質問お願いします
質問⑤ Kiyoshi Miyata
僕もこちら興味本位の質問ですが、光で局在を制御するときに、どのくらいの空間分解能を出せるるとよさそうなんでしょうか?
質問⑥ moriken55
なぜ線虫では使えるのに、マウスでは使えないのでしょうか。血中滞留性が良くないので、標的組織に到達できなにでしょうか。あるいは、血中でペプチド部が分解するのでしょうか。
質問⑦ moriken55
通常のリガンド添加によるシグナリングに比べて、SLIPT法のシグナリングは強度や質に違いはあるでしょうか。
質問⑧ moriken55
なるほどアルキルをケージするとか?
質問⑨ Akihisa Mino
細胞集団の中でRasやTiam1等での細胞活性化の細胞の数を段階的に増やす実験を行うと、がん細胞の転移浸潤開始の細胞数の閾値を検討することも可能になりますでしょうか。
コメント❶ moriken55
面白いです! 不思議~
質問⑩ Hiroshi Kamizawa
アルブミンにトラップされるとのことでしたが、アルキルの長さを変更することでアルブミンへの親和性を低下させることは可能でしょうか?そもそもアルキル部分の影響でトラップされているのでしょうか?
コメント❷ moriken55
細胞外に来てくださいよ。 学生用と聞いたもので
